1. 発表者:

柳下 祥 東京大学大学院医学系研究科附属疾患生命工学センター 構造生理学部門/
東京大学国際高等研究所ニューロインテリジェンス国際研究機構(WPI-IRCN)講師
飯野 祐介 東京大学大学院医学系研究科附属疾患生命工学センター 構造生理学部門/
東京大学国際高等研究所ニューロインテリジェンス国際研究機構(WPI-IRCN)研究員
河西 春郎 東京大学大学院医学系研究科附属疾患生命工学センター 構造生理学部門 教授/
東京大学国際高等研究所ニューロインテリジェンス国際研究機構(WPI-IRCN)主任研究者
2. 発表のポイント:
◆統合失調症治療薬(抗精神病薬)として広く使われているドーパミン2型受容体(D2受容体)の阻害剤が幻覚や妄想などの症状を改善する仕組みの理解を進めました。
◆報酬を予測する音を区別する学習課題を行うマウスにおいて脳内光刺激を用い、D2受容体はドーパミン濃度の低下を検出して過剰な学習を現実に合わせて訂正する弁別学習を起こし、この機構の障害は抗精神病薬により回復することを見出しました。
◆脳が環境に適応する新原理と、精神症状の発症原因にシナプスの障害の可能性があることを提案し、統合失調症の早期診断に新しい道を拓きました。
3.発表概要:
脳内報酬物質といわれるドーパミン(注1)の2型受容体(D2受容体)は抗精神病薬の主要な標的ですが、シナプスや行動の制御機序は不明でした。東京大学大学院医学系研究科・IRCNの飯野祐介研究員、澤田健大学院生、山口健治研究員、河西春郎教授、柳下 祥講師およびIRCN・京都大学工学部情報学研究科の石井信教授らのグループは、マウスの行動実験において光による神経活動の操作・観察技術を組み合わせることで、環境情報から報酬を予測する記憶が間違っていた際に側坐核(注2)で生じるドーパミンの一過性低下をD2受容体発現細胞が検出し、間違った記憶を訂正していることを発見しました。さらにマウスの脳スライスを光操作で観察したところ、微小なドーパミン信号変化をD2受容体が検出しスパイン(注3)が頭部増大(注4)を起こすことを見いだしました。この鋭敏な検出機構は覚せい剤によりドーパミン濃度がわずかに亢進すると破綻しますが、D2受容体阻害薬(抗精神病薬)により回復しました。
本研究は脳の新しい学習原理を明らかにし、統合失調症などにおける精神病症状を説明する新しいシナプス仮説を導きました。
本研究は、文部科学省科学研究費助成事業 、IRCN、日本医療研究開発機構(AMED)、JST戦略的創造研究推進事業(CREST)の支援を受けて、国際科学誌「Nature(電子版)」に2020年3月18日付オンライン版で発表されます。
4.発表内容:
【研究の背景】
統合失調症では神経伝達物質のドーパミンが過剰になると妄想・幻覚などの症状を生み、D2受容体を抗精神病薬が遮断すると症状が改善することが知られています。このようにD2受容体は脳機能において重要な役割を担いますが、具体的にどのようにシナプス機能や行動を制御しているのかは未解明でした。
ドーパミン神経が強く投射する領域である側坐核にはドーパミン1型受容体(D1受容体)を発現する神経細胞群(D1細胞)とD2受容体を発現する細胞群(D2細胞)が半数ずつ存在します。報酬による条件づけ学習(注5)において予想外の報酬が与えられると、定常状態から一過性にドーパミン濃度が上昇します。これにより音などの条件刺激(CS)と水などの報酬(無条件刺激, US)を連合させる古典的条件づけ学習が生じます。この結果、学習前は反応を起こさなかったCSから動物は報酬を予測し、条件反射が起きるなど、CSが高い価値情報を持つようになります。当研究室では以前、ドーパミンの一過性上昇をD1細胞のスパインが検出し頭部増大を起こすことが、報酬による条件づけ学習を制御する基盤となることを報告しました。一方、ドーパミン濃度は予想外に報酬が来ないと一過性に濃度が低下することが知られています。このドーパミンの一過性低下をD2受容体が検出することが予想されていましたが、その実態の検証は電気刺激法などの古典的な実験手法では困難でした。
しかし、光遺伝学(注6)の登場によりドーパミン濃度を脳スライス上で厳密にコントロールすることが可能になりました。この方法と当研究室で開発したグルタミン酸2光子アンケージング(注7)を組み合わせることで、単一スパインレベルでD2受容体がドーパミン濃度変化によりシナプス機能を制御する機序の研究が可能になりました。また、このようなシナプスレベルのドーパミン検出機序が実際の学習行動とどのように関係するのかについても光遺伝学(ドーパミン光操作)や高感度の神経活動センサーと光ファイバーを介した蛍光検出(ドーパミン光測定)を組み合わせることで検証可能になりました。そこで、光による神経活動の操作・観察技術を駆使し、脳スライスと学習行動の両方を調べることで、D2受容体機能の謎に迫りました。
【研究内容】
マウスを用いて音(CS)と報酬を連合させる条件づけ学習を調べたところ、報酬による条件づけ学習は実際に提示した低周波数の音(CS+)だけでなく、提示していない高周波数の音にまでに広がる(汎化)ことが分かりました(図1A)。この汎化する学習にはD1受容体が必要で、視覚刺激にまでは広がりませんでした。現実世界において食べ物や水など報酬を予測する手がかり情報にはある程度のばらつきがあるため、汎化学習は報酬を効果的に獲得するのに合理的で、進化の過程でこのような回路が生得的に備わったと考えられます。
次に、条件づけに用いたCS+を報酬ありとし、高周波数の音(CS-)を報酬なしとすることを繰り返すと、最初は汎化的にCS-に対して見られた条件反射は2日間の学習で減少しました(図1A)。この弁別学習(注8)の最中に、側坐核でドーパミン神経活動の光測定を行うと、予測した報酬が実際にはもらえないCS-の提示後に2秒程度ドーパミンが低下していました。さらにこのドーパミンの一過性低下を光による神経活動操作により打ち消すと弁別学習ができなくなったことから(図1B、 図2A)、ドーパミン一過性低下は予想した報酬が実際にはない場合にがっかり信号として予測を精緻化すると考えられました。一方、全ての刺激から報酬をなくす消去学習(注9)ではドーパミン一過性低下は起きませんでした。
そこで、マウスの脳スライスを用いてこのドーパミン一過性低下を検出する仕組みを詳しく調べました。まず2光子励起によりD2細胞の単一スパインをグルタミン酸刺激することで、スパイン頭部増大が起こる条件を見いだしました。次に、光遺伝学を駆使してドーパミン神経を青色光で刺激し、脳スライス上に生体で見られる定常状態のドーパミン活動を再現しました。この定常条件ではD2受容体がグルタミン酸刺激によるスパイン頭部増大を抑制していましたが、ドーパミン刺激を0.4秒停止してドーパミン濃度を低下させると、スパイン頭部増大が起こりました(図2B)。この頭部増大に必要な信号をマウス個体において阻害すると弁別学習ができなくなりました。脳スライスと学習行動の結果を合わせると、ドーパミン一過性低下によるがっかり信号をD2細胞のスパインが、D2受容体を介して検出することで過剰な予測を訂正すると考えられます(図1B)。
また、覚せい剤は脳内のドーパミンを増やし、ヒトにおいて幻覚や妄想といった精神病症状を起こすことがあります。覚せい剤投与時のドーパミン動態を調べると、定常状態のドーパミン濃度が増加し、ドーパミン一過性低下信号が小さくなっていました。この状態ではD2細胞のスパイン増大および過剰な予測を訂正する学習が障害されました。この障害はD2受容体遮断薬(抗精神病薬)により回復しました。
【社会的意義・今後の展望】
本研究の結果により、報酬学習を行う際にまずD1細胞が汎化的に学習し、その後、D2細胞が現実に合わせて訂正する弁別学習を行うという新しい脳の計算原理が見いだされ(図1B)、人工知能研究への応用が考えられます。
さらにD2細胞が過剰な予測を訂正する機序はドーパミン過多によって障害され抗精神病薬により回復することから、この機序の破綻が統合失調症初期の妄想を認知心理学的に説明する価値情報の帰属障害(注10)を引き起こす可能性があります(図1C、D)。また、消去学習は価値に関する状況処理を脳が切り替えることで生じ、D1/ D2細胞は価値の状況ごとに汎化・弁別学習をすると考えられます。
これらのことから、就学、就職、移民などの新しい価値が関わる生活状況において汎化・弁別をする際に、D2受容体の機能の脆弱性などのシナプスの異常があると、弁別学習障害を介して妄想の発展に至るという、分子、シナプス、回路、個体行動、リアル・ワールドまで整合性のある新しい仮説が提起でき、病態理解や早期診断に進歩をもたらすと期待されます。
5.発表雑誌:
雑誌名:Nature(3月18日オンライン版)
論文タイトル:Dopamine D2 receptors in discrimination learning and spine enlargement
著者:Yusuke Iino*, Takeshi Sawada*, Kenji Yamaguchi*, Mio Tajiri, Shin Ishii, Haruo Kasai† and Sho Yagishita†
(*共同第一著者、†共同責任著者)
DOI番号:https://doi.org/10.1038/s41586-020-2115-1
アブストラクトURL:https://www.nature.com/articles/s41586-020-2115-1
6.問い合わせ先:
<研究内容に関すること>
東京大学大学院医学系研究科 附属疾患生命工学センター 構造生理学部門
教授 河西 春郎(かさい はるお)
東京大学大学院医学系研究科 附属疾患生命工学センター 構造生理学部門
講師 柳下 祥(やぎした しょう)
<JST事業に関すること>
科学技術振興機構 戦略研究推進部 ライフイノベーショングループ
川口 哲(かわぐち てつ)
E-mail:crest@jst.go.jp
7.用語解説:
(注1)ドーパミン
中脳にあるドーパミン神経から放出される神経伝達物質で意欲と報酬による学習など制御する。ドーパミン神経は生体では5Hz程度の周波数で活動しており、脳内には常に一定量のドーパミンが存在し、報酬や罰など外的な刺激により濃度が変化する。
(注2)側坐核
側坐核は新皮質や海馬、扁桃体といった脳領域からグルタミン酸作動性の興奮性入力を受けると同時に腹側被蓋野からドーパミン神経の入力を受ける。ドーパミン神経がグルタミン酸神経伝達の可塑性を調節することで報酬による学習が起こると考えられている。
(注3)スパイン
神経細胞間の接合部位をシナプスと呼び、シナプス前部からグルタミン酸が放出され、シナプス後部で受容されることで神経活動が伝達される。スパインはグルタミン酸作動性シナプスの後部に存在する1ミクロン以下の棘状の構造物であり、大きいスパインほどグルタミン酸感受性が高い。スパインという突出構造はシナプスの個別可変性を起こし記憶の集積性を高める。スパインはさまざまな種の動物の脳の最も高次中枢の神経によく発達している。その形は多形であり、その頭部の大きさがシナプス結合強度を決める。
(注4)頭部増大
スパイン頭部の体積が増大することで表面受容体の数が増えシナプス結合強度が増加する。このようにシナプス結合強度が変化することをシナプス可塑性と呼ぶ。スパイン頭部増大の様な形態可塑性は速く起き長期化するので学習・記憶との関連が強い。
(注5)条件づけ学習
動物に無条件に反応を引き起こす刺激(無条件刺激、例えば水の提示は舐める反応(リック反応)を引き起こす)を特に反応を起こさない刺激(条件刺激、CS、例えば音)の後に繰り返し提示すると、条件刺激だけで動物は反応を起こす(条件反射)ようになる。このような学習を古典的(パブロフ)条件づけと呼ぶ。
(注6)光遺伝学
光活性化タンパク質を細胞に遺伝子導入することで、細胞機能を光により制御する技術。本研究
においては光の照射により細胞内に陽イオンを流入させることで細胞を発火させることができるタンパク質をドーパミン神経細胞に発現させ、光によるドーパミン神経細胞の操作を行った。
(注7)2 光子アンケージング法
2 つの光子が同時に分子に吸収される現象を2光子励起という。対物レンズにより超短パルスレーザーを焦点に集約させることで、微小な体積でのみ2光子励起を誘発することができる。この方法をケージド・グルタミン酸(不活性化されたグルタミン酸)に応用することで、微小体積でグルタミン酸が活性化され、従来の電気刺激では不可能だった単一のスパインへのグルタミン酸刺激が可能となった。この励起は赤外光を使うので、光遺伝学と併用することができる。
(注8)弁別学習
2つ以上の知覚刺激を用いて、それぞれの知覚刺激に対して異なる反応が生じるようになる学習のことを弁別学習という。本研究においては一方の刺激に対しては報酬を与え、他方の刺激には報酬を与えないことで、報酬の有無を弁別して動物が条件反射を示すようになる古典的(パブロフ)条件づけの弁別学習を用いた。この報酬の有無を弁別する学習は、例えば聴覚皮質が関わるとされている音の高低そのものの弁別学習(音の高低を弁別して異なる行動をとることで報酬を獲得する学習)とは異なると考えられている。
(注9)消去学習
条件づけ学習後に、条件刺激(CS)のみを無条件刺激なしで与えることを繰り返すと、条件反射が起こらなくなっていくこと。弁別学習では報酬がもらえるCSを区別することを学習するが、消去学習では報酬が全く来なくなったことを学習する。
(注10)価値情報の帰属障害
統合失調症では何らかの原因により一般には報酬や罰などの価値を伴わない環境情報に対しても価値を見いだすことが妄想などの精神病症状の基盤にあるものと考えられるようになってきた (Kapur, S. American journal of Psychiatry, 2003)。
8.添付資料:
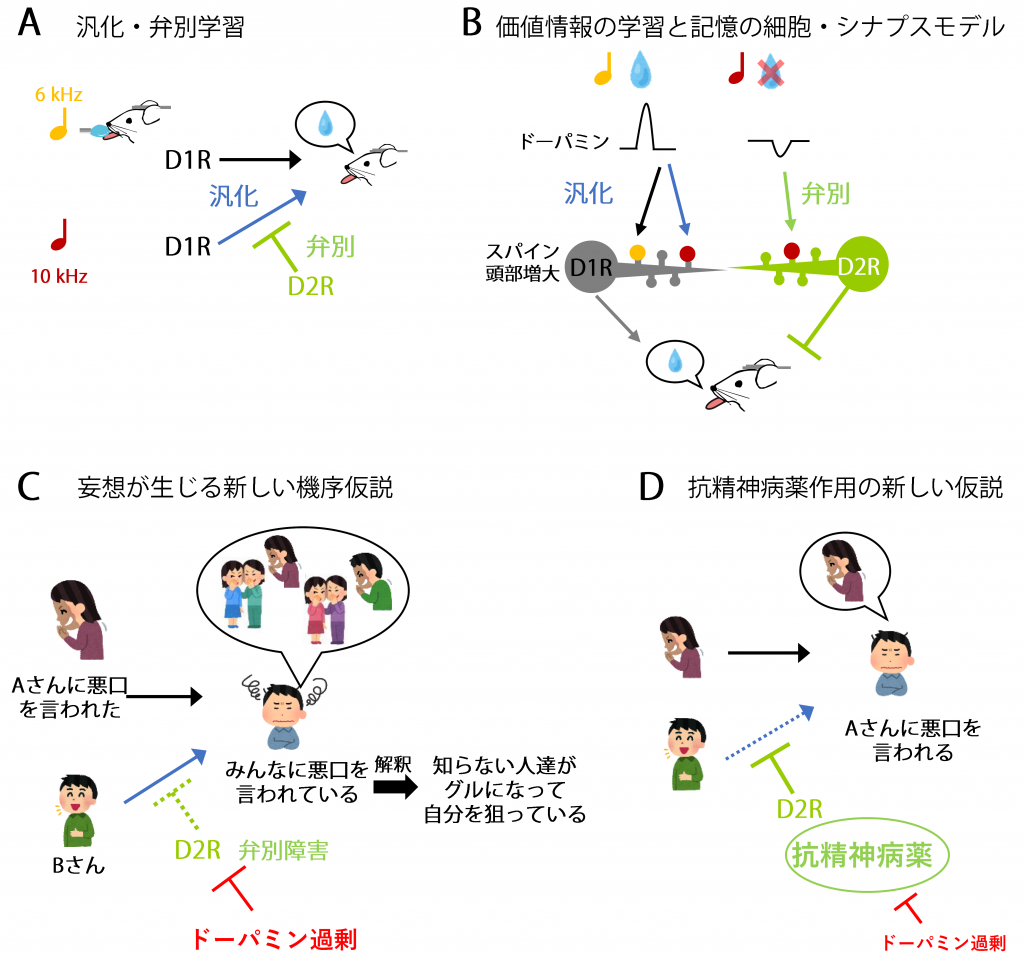
図1.汎化・弁別学習のモデルと新たな精神病症状の仮説
A. 汎化・弁別学習。古典的条件づけ学習において6 kHzの音の提示の後にマウスに報酬を与えると、D1受容体を介して実際に聞いた6 kHzのみならず、聞いていない類似の知覚情報(10 kHzの音)にまで汎化して報酬予測をする(青矢印)。しかし、この汎化した価値の予測が誤っていた際にD2受容体を介して誤った価値記憶を抑制する弁別学習が起こる(黄緑線)。
B. 価値情報の記憶学習の細胞・シナプスモデル。報酬はドーパミンの一過性上昇を起こし、D1受容体発現細胞のグルタミン酸シナプス・スパインの頭部増大を起こす。このシナプス可塑性が実際に経験した知覚入力(6 kHzの音)に加えて、類似の知覚入力(10 kHzの音)にまで広がった価値情報の記憶を保持する(青矢印)と考えられる。しかし、予想した報酬が実際には来なかった場合には、ドーパミン一過性低下が生じ、D2受容体発現細胞のスパインの頭部増大が起こる。これにより、誤った価値情報の連合記憶を抑制する弁別記憶が形成される(黄緑線)。D1細胞とD2細胞が協調して働くことで、環境の価値情報を脳内に素早く適切に表現し、行動するものと考えられる。
C. 妄想が生じる新しい機序仮説。統合失調症では周囲の皆が悪口を言っているのではないかなどと感じてしまう被害妄想があり、その原因として周囲の環境情報に対して過剰な価値情報を帰属してしまうことが考えられている。本研究では報酬を用いたが、不快な刺激によってもドーパミンが上昇する部位があることが分かっており、今回の研究成果から下記のような新しい妄想発症機序のモデルが考えられる。実際にAさんに自分の悪口を言われ不快に思うとドーパミンが上昇し、D1受容体を介してAさんの話をしている姿に対して不快な価値情報が割り当てられ、同時に汎化して他の人に対しても不快な価値情報が広がって割り当てられる。通常、Bさんは悪口を言っていないということから自分の価値情報を訂正できるが、ドーパミン過剰状態などでD2受容体経路がうまく働かないと誤った記憶の訂正ができなくなる。この結果、現実と乖離した価値情報の帰属障害が起こる。この状態が悪化すると次第に自分は狙われているのではないか、監視されているのではないかというように認知的に解釈され、妄想が発展するものと考えられている。
D. 抗精神病薬はD2R受容体に作用して弁別学習機能を回復させることで、現実と乖離した認知を訂正すると考えられる。このように経験とシナプス過程を介して妄想が生じることを説明しえる新しいモデルが導かれた。
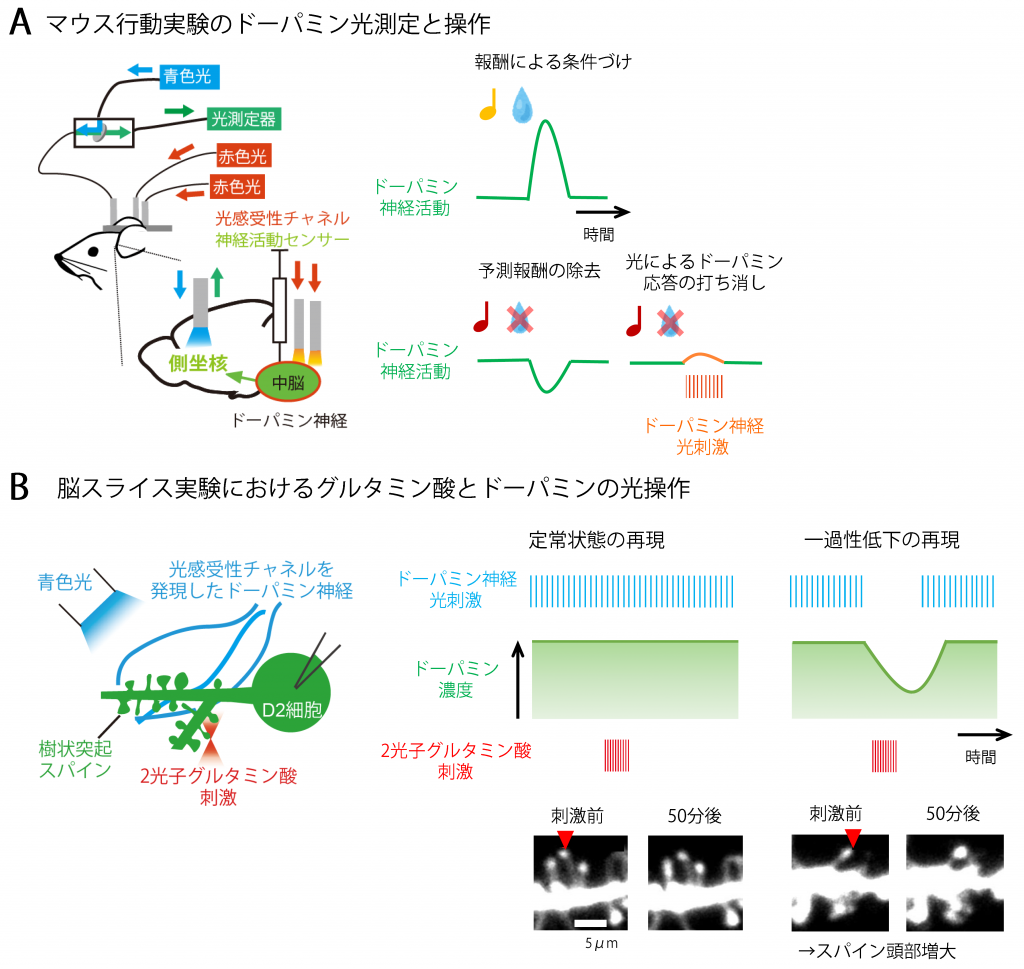
図2.光を駆使したマウス行動実験とスライス実験によるD2受容体機能の解明
A. 光によるドーパミン神経活動の観察と操作を組み合わせたマウス行動実験。青色光で励起すると神経細胞の活動に応じて緑色蛍光を示すセンサー・タンパク質と赤色刺激によって神経細胞を発火させることができるチャネル・タンパク質を、ウィルスベクターを用いてドーパミン神経に導入した。光ファイバーをマウス脳に留置し緑色蛍光でドーパミン神経活動を観察すると、条件づけ学習の際に報酬の提示により一過性にドーパミン神経活動が上昇していた。一方、弁別学習の際に予想した報酬がやってこなかった時、ドーパミンは2秒程度の間だけ活動が低下していた。赤色光刺激によりドーパミン神経を僅かに活性化させて一過性低下の信号を打ち消すと、マウスは弁別学習ができなくなったことから、ドーパミン神経活動と行動の関係を明らかにした。
B. 光によりドーパミンとグルタミン酸を操作する脳スライス実験。光感受性チャネル・タンパク質をドーパミン神経に導入したマウスの脳スライスを作成し、青色光でドーパミン神経を活性化させることができるようにした。また赤外光による2光子励起によりグルタミン酸刺激を単一スパインに与えることができる。この実験系においてドーパミン神経を生体の活動である5 Hzで刺激すると、定常状態のドーパミン濃度を再現することができた。このドーパミンはD2受容体を介して、グルタミン酸刺激によるスパイン可塑性を抑制した。しかし、弁別学習の際に見られたドーパミンの低下があると、グルタミン酸刺激による頭部増大が見られた。このことからドーパミン一過性低下の僅かな信号を検出するのにD2受容体を介したスパイン機序が重要と考えられた。


